4. Принцип устройства
обучаемых систем управления
Биогидравлическая модель нервной системы. В основу принципиального устройства обучаемых систем управления положено техногенное представление о мозге животных, точнее говоря — биогидравлическая модель нервных систем живых существ, поэтому обучаемые системы можно считать техническими нервными системами. Биогидравлическая модель, о которой идет речь, оказалась достаточно плодотворной для того, чтобы на её основе создать работоспособную систему управления.
В истории
нейрофизиологии хорошо известны многие инженерные концепции, и ни одна из них
не смогла отразить с достаточной степенью соответствия человеческий мозг,
однако некоторые из них были использованы
продуктивно в самой технике. Достаточно вспомнить компьютерную и перцептронную
модель, согласно которым мозг представляет собой вычислительный процессор с
чрезвычайно объёмным блоком
документальной памяти, либо многослойную систему электрических проводников с
перекрещивающимися случайными связями. Первая модель породила электронную
вычислительную технику, вторая —
нейронные сети, постепенно завоёвывающими признание не только в науке, но и в
технике. Неприятие техногенных концепций специалистами-нейрофизиологами не
может восприниматься как запрет на иной,
небиологический поход к работе мозга и всей нервной системы в целом, поскольку
отдача от него может возникнуть совсем не в области биологии, а в иной сфере
Биогидравлическая
модель нервных систем проповедовалась в своё время великим французским ученым
Декартом (1596-1650), а еще раньше —
всемирно известным греческим мыслителем Аристотелем (384-322 до н.э.). Но не только авторитеты заставили возвратиться
к указанной модели. Гидравлика, как вид управляющих потоков, была оттеснена в нейрофизиологии с момента первой
регистрации электрических биотоков, существование которых оспорить уже было
невозможно. Всеобщее увлечение электричеством захватило нейрофизиологию
настолько, что все прочие виды энергии попросту были вычеркнуты из числа возможных
в нервных системах» И нужно было проявить определенную волю, чтобы заставить
нейрофизиологов принять, например, химическую концепцию синапса, и сам этот
факт стал крупным событием. О гидравлике же не вспоминают даже тогда, когда говорят
о функционировании жизнеобеспечения нервных клеток. В то же время целый ряд специфических факторов в работе нервных
систем и мозга в том числе свидетельствует в пользу биогидравлической модели.
Так, на поведение животных и человека со всей очевидностью влияет изменение атмосферного давления. Или — на внутриклеточном уровне: скорость
перемещения так называемого потенциала действия больше соответствует скорости
распространения волны давления в вязкой жидкости
и совсем не соответствует скорости электричества; в некоторых (коротких)
нервных клетках электрический потенциал действия не регистрируется, то есть
отсутствует вообще; только эластичные гидравлические каналы, какими являются
каналы нервных клеток, нуждаются в ужесточении их оболочек миелином, а
электричеству миелин ни к чему; указанное ужесточение способствует повышению
скорости распространения потенциала действия, то есть волны давления;
разрушение миелиновых оболочек, как форма болезни, приводит к нестабильности, в
движениях человека, равно такой же, какая
возникает в работе гидравлических механизмов с длинными эластичными
слабоармированными шлангами; скорость перемещения импульсов по нейронам прямо
зависит от диаметра аксона, и даже такой
общеизвестный факт, как вздрагивание ноги при ударе молоточком по колену,
говорит в пользу гидравлических процессов.
Биогидравлическая модель не отвергает наличие электрических биотоков, но отводит им иную роль: они необходимы для поддержания волны давления и для воздействия на мышцы (но не принимают участия в возникновении возбуждений рецепторов, в выработке сигналов управления и не служат средством передачи этих сигналов). Не отвергает она также и биохимические процессы, напротив даже — опирается на них: гидравлическая среда — биологически активна; кроме функции формирования и передачи сигналов управления она еще обеспечивает по совместительству питание нервных клеток и способствует изменению проходных сечений синаптических щелей. Другими словами, биогидравлика решает все задачи жизнедеятельности нервных систем в комплексе, а именно:
· чем больше поток жидкости, тем больше сигнал управления и тем энергичнее движение соответствующей мышцы;
· чем больше поток жидкости вообще, тем здоровее нервная клетка и тем больше скорость её роста (если она ещё растет);
· и, наконец, если поток жидкости исчезает в результате падения давления до нуля, например в результате разрыва нерва, то парализуется движение и возникает ощущение боли как свидетельство приостановки местной жизнедеятельности организма: нет управляющего потока — нет и жизни.
Поэтапность раскрытия тайны мозга. Для
того, чтобы характеризовать нервные системы в целом, не достаточно одной биогидравлической
модели, отражающей только природу управляющей энергии, — необходимо ещё определить систему связей нервных клеток; и здесь
уместно воспользоваться подсказкой нейрофизиологов, утверждавщих, что, хотя
нервная система есть управляющее устройство, но механизм управления унаследован ею от тех управляющих систем, которые существовали
в живой природе задолго до возникновения мозга
/1.7/. Намек можно расшифровать так: сначала нужно понять организацию
нейронных систем простейших существ, руководствующихся исключительно
безусловными рефлексами, то есть таких систем, которые не способны ни
запоминать и ни забывать; освоив этот рубеж, можно было переходить к
следующему, имея в виду, что для реализации запоминания-забывания нужно ввести
в нейронные системы некоторое, вполне определённое,
конструктивное (неудачный термин для биологии) изменение, и таким изменением
является, скорее всего, введение синаптической щели с изменяющейся проводимостью,
и нужно определить факторы, влияющие
отдельно на увеличение и отдельно на уменьшение указанной проводимости, то есть
нужно хорошо освоить рубеж условных рефлексов сначала с обучением при внешней
стимуляции, то есть с обучателем, а затем и без него — в виде самообучения на основе игры и подражания о критериями
«хорошо» и «плохо»: и только после этого можно попытаться приступить к
раскрытию тайны мозга человека, несомненно
имеющего своё конструктивное отличие от мозга животных, такое отличие, которое
подарило человеку речь и ту волю, с помощью которой он может ставить себе цели
и принуждать себя на переключение с решения одной задачи на другую. (Перед
последним рубежом стоило бы задуматься: а одолевать ли его вообще или обождать
до лучших времен?). С нейрофизиологами можно согласиться в том смысле, что
предложенный ими путь поэтапного освоения может оказаться во времени куда
короче, чем штурм той неприступной скалы, которую представляет собой в
настоящее время мозг человека.
Обучаемые системы
управления соответствуют нервным системам, включающим мозг, тех живых существ,
которые способны вырабатывать навыки, и
на большее они не претендуют. Они могут осуществить поиск, но не способны
распознавать, и эти два действия нельзя смешивать. Любое животное способно
осуществлять поиск — можно даже утверждать,
что поиск является основной, чуть ли не единственной, функцией деятельности
животных, — но распознавать, то есть классифицировать и сортировать, может
только человек. Распознавание в строгом смысле не является навыком; это — продукт логики, то есть особой способности,
характерной исключительно для мозга человека. Достаточно человеку
повнимательнее присмотреться к самому себе, и он отметит явное различие в таких
своих действиях, как, например, обычная ходьба и переставление ног по подсказке
или по инструкции, как машинальное
написание слова и выведение букв на ранних стадиях школьного обучения, и т.д.
Первое — навык, второе — логическое
действие; первое вырабатывается в результате многократных
повторов при обучении, второе выполняется по командам, следующим одна за
другой в определенной логической последовательности. Так, большую печатную
букву А дети истолковывают как имеющую вид шалашика с перекладиной
посредине, а букву Н — как два столбика с той же перекладиной и
т.д. Позднее, когда вырабатывается автоматизм написания или прочтения, отпадет
необходимость в подобном толкований; автоматизм и есть навык. Курица клюёт
червя автоматически, нисколько не задумываясь о том, похож или не похож он на
ржавый гвоздь, а цыпленок, недостаточно обученный, может попытаться склевать и
его, но и он — цыплёнок — распознаванием
не занимается: просто его навык ещё не до конца сформировался.
В отличие от обучаемых систем управления перцептрон и развивающиеся на его основе нейронные сети создаются не для выработки навыков, а для решения задач распознавания. Будут ли они полным подобием человеческого мозга — покажет будущее (скорее всего — нет), но то, что они способны распознавать — не вызывает сомнения уже сейчас, и их распознавание отличается от компьютерного тем, что решение вырабатывается не путём расчёта по решающим правилам, а в результате обучения. Впрочем, распознавание может быть использовано, также как и навык, для управления исполнительными органами любого объекта, но только в результате многоходовых действий: сначала по определённым правилам надо перевести внешний образ ситуации во внутренний образ очувствления; затем следует подобрать из имеющихся в памяти образов тот, который меньше других отличается от полученного; после этого надо извлечь из памяти соответствующий комплект сигналов управления, и, наконец, остается распределить по соответствующим исполнительным органам. Логика здесь очевидна, но также очевидно и то, что мозг животного работает иначе.
Итак, первый
рубеж — безусловные рефлексы простейших живых существ. Им соответствуют
простейшие нейронные системы, реализующие простые законы управления, известные
в нейрофизиологии как «стимул-ответ». В технике им соответствуют логические
автоматические системы управления со всевозможными датчиками, в которых всякому
состоянию датчиков соответствует однозначный ответ исполнительных органов. В
качестве примера можно привести станочный автомат (или полуавтомат) с концевыми
переключателями, у которого порядок взаимосогласования положений переключателей
(датчиков) и работы двигателей определен его циклограммой.
Появившиеся на заре биологической истории безусловные рефлексы не были устранены на последующих этапах развития живых существ, а передавались как ценное свойство от одного вида к другому, и так дошли они до человека. Из простых его безусловных рефлексов можно назвать отдергивание руки при болезненном уколе пальца, из более сложных — аккомодация глаз, то есть автоматическое наведение их на резкость.
Второй рубеж — навыки и условные рефлексы. Этот рубеж наукой, мягко говоря, ещё не освоен. Сами нейрофизиологи не скрывают того факта, что они пока не могут предложить ни одной удовлетворительной концепции, раскрывающей как единое целое устройство, обучение и работу нервных систем животных и тем более человека. Сегодня нейробиология, образно говоря, блуждает в дебрях эмпирики, накопившейся за многие годы практических исследований мозга, и не может найти той тропы, которая ведёт к раскрытию его тайны.
Такое состояние сдерживает развитие техники. Существующие системы управления вплоть до компьютерных используют пока почти исключительно параметрическую информацию, но ведь кроме неё существует еще признаковая, факторная и, особо выделим, образная: зрительная, слуховая, тактильная и другие, — информации, И все эти «прочие» виды информации практически пока не задействованы. Может быть именно поэтому роботы, даже самые совершенные, не нашли себе широкого применения, и мешает этому их слепота, глухота, безчувствительность. Дело усугубляется ещё и тем, что логическое переваривание собранной с помощью датчиков информации имеет свои пределы, ведь логику — так называемое математическое обеспечение — формирует человек, а его возможности ограничены.
Техническая модель нервных систем. В такой обстановке инженерам не оставалось ничего другого, как самим предлагать технические системы, реализующие навыки и условные рефлексы. Одной из таких систем и явилась обучаемая система управления, создание которой стало результатом нового инженерного подхода к нервным системам животных. В принципе задача раскрытия секрета нервных систем ставилась так: живой организм имеет рецепторную оболочку — сетчатку глаз, ушные рецепторы, чувствительную кожу и прочее, — вырабатывающую информацию очувствления; имеет нейронную сеть, поставляющую эту информацию в мозг; имеет кору головного мозга, преобразующую информацию в сигналы управления, и, наконец, имеет систему мотонейронов, доставляющих эти сигналы управления к мышцам, — и нужно было ответить на следующие вопросы: как рецептор вырабатывает свою информацию? как передается эта информация по нейронам? как распределяется эта информация между нейронами? что такое синапс? Какова особенность синапса коры головного мозга? имея в виду, что именно эта особенность определяет в конце концов законы управления? как вырабатываются в коре головного мозга сигналы в конце концов, сигналы управления доставляются к мышцам?
Ответы будут
сугубо инженерными и построены они, скорее всего, по принципу не «как есть», а «как надо», то есть решается задача
не раскрытия биологической тайны, а конструирования технического секрета
навыков и условных рефлексов.
Начнём отвечать по порядку. В рамках биогидравлической модели рецептор представляет собой колбочку, заполненную управляющей жидкостью и соединённую с каналом нейрона (рис. 1.7).

Рис.1.7.
Гидравлическая модель системы навыков и условных
рефлексов
При
воздействии на колбочку внешнего раздражителя часть жидкости выдавливается в
канал; в качестве раздражителя могут выступать: свет, касание, давление,
химическое воздействие и прочее. Реакция рецептора на внешнее воздействие может
быть импульсной, и тогда на выходе его последует череда импульсов в виде волн давления. Для удобства импульсы можно
суммировать и представлять в форме аналогового непрерывного сигнала.
Усреднённое избыточное давление управляющей жидкости, создаваемое i-ым рецептором на выходе под воздействием внешнего
раздражителя, договоримся называть возбуждением рецептора и обозначим через ![]() . Договоримся также под рецептором понимать только
элемент очувствления и ничего больше.
. Договоримся также под рецептором понимать только
элемент очувствления и ничего больше.
Рецепторы бывают двух типов: прямого действия и обратного. Например, фоторецептор прямого действия возбуждается на свету, а фоторецептор обратного действия — в темноте. Количество тех и других — одинаковое, так что на один рецептор света приходится один рецептор темноты, а вместе они составляют рецепторную пару. Этим можно объяснить экспериментально установленное наличие возбуждающих рецепторов, увеличивающих результирующий сигнал управления при воздействии на них раздражителя, и тормозящих, уменьшающих тот же сигнал в тех же условиях. Однако, если говорить не об отдельных рецепторах, а о парах, то разделение их на возбуждающие и тормозящие происходит в процессе обучения и будет подробно рассмотрено ниже.
Возбуждения
рецепторов ![]() могут быть
усилены или ослаблены эмоциями. С инженерной точки зрения эмоции проявляются в
виде изменения напряжения той среды, в которой располагаются рецепторы, или в виде изменения давления крови. В
электрических обучаемых системах управления возбуждения рецепторов можно
изменить путем изменения напряжения их общего питания.
могут быть
усилены или ослаблены эмоциями. С инженерной точки зрения эмоции проявляются в
виде изменения напряжения той среды, в которой располагаются рецепторы, или в виде изменения давления крови. В
электрических обучаемых системах управления возбуждения рецепторов можно
изменить путем изменения напряжения их общего питания.
Представив
рецептор в виде колбочки с жидкостью и с одним отходящим каналом, нужно
ответить на вопросы: на какое время хватает жидкости и как она там снова
накапливается? — ведь нервная система человека не закольцована как кровеносная.
Ответ по времени нужно искать в суточном цикле: запасов жидкости в рецепторе,
очевидно, хватает в среднем на день, на время бодрствования; после чего давление внутри рецептора падает
настолько, что он нуждается в подзарядке; и тогда наступает сон. Таким
образом, сон есть не что иное, как состояние организма, в котором
осуществляется подзарядка рецепторов; их
возбуждения снижаются ниже порогового уровня. Засыпают, то есть переключаются
на подзарядку, все органы чувств не одновременно; также не одновременно они
просыпаются. То время, когда спят все
рецепторы, характеризуется как глубокий сон; мышление человека при частично
отключенных (спящих) органах чувств — ущербно, дефектно и называется
сновидением.
Нейрон,
согласно той же биогидравлической модели, представляет собой микротрубочку, по
которой управляющая жидкость направляется в мозг. Каждая такая микротрубочка
полностью изолирована от других и нигде с ними не соединяется: она несёт свой
сигнал, то есть возбуждение соответствующего рецептора, неизменным до самой коры головного мозга. Несколько микротрубочек
могут быть собраны для компактности в пучок —
это и есть аксон многоканального нейрона, а на конце пучка они разветвляются и
представляют собой аксонные окончания.
Если длина микротрубочки не достаточна для соединения рецептора с мозгом, то
составляется цепочка из микротрубочек. Соединение таких микротрубочек
осуществляется с помощью нерегулируемых
синапсов, то есть особых точек, в которых окончания одного многоканального
нейрона присоединяются к дендритам или к телу другого. Принцип независимости
(автономности) отдельных информационных
каналов, какими являются микротрубочки, распространяется также и на вою
цепочку. Микротрубочки эластичны, и это свойство, как известно из гидравлики,
может сделать неработоспособной любую гидросистему: нежёсткость трубопроводов
приводит к неопределённости поток жидкости и к снижению их скоростей. Поэтому
пучек микротрубочек в виде аксона имеет миелиновую оболочку, ужесточающую
каналы. Говоря о передаче информации от рецептора к мозгу, стоит ещё раз
повторить, что в принятой модели нервных систем животных каналы, по которым распространяется
эта информация, независимы и нигде друг с
другом не соединяются. Такое утверждение приходится принимать на веру, зная,
что оно противоречит современным представлениям нейрофизиологии, хотя та же нейрофизиология однозначно утверждает, что
конкретной группе рецепторов в мозгу соответствует конкретный участок коры.
Определимся с терминологией: и отдельную микротрубочку, и непрерывную цепочку
микротрубочек (там, где она есть), передающих сигнал от рецептора к коре головного
мозга, договоримся называть просто нейроном, а канал, идущий от мозга к мышцам
— мотонейроном.
У последнего функции несколько шире, чем у нейрона; в коре мотонейрон собирает управляющую жидкость от всех рецепторов очувствления, затем транспортирует её к определённой мышце (мышечному волокну) и воздействует на неё, побуждая к сокращению; сила воздействия пропорциональна управляющему потоку. На мотонейроны распространяется тот же самый принцип независимости: они нигде не соединяются и не разветвляются. Дендритность, которую постоянно демонстрирует нейроанатомия, не должна нас смущать: мотонейроны, как отдельные каналы, могут быть собраны, как и нейроны, в пучок, образуя многоканальные мотонейроны со многими окончаниями аксона, но соединений внутри пучка нет. Это утверждение (а точнее — предположение) не исключает соединений управляющих каналов нейронов и мотонейронов в принципе. Они возможны, например, в целях дублирования, когда несколько рецепторов «работают» на один нейрон или когда несколько мотонейронов (точнее — несколько каналов мотонейрона) подсоединяются к одной мышце; такую множественность можно проигнорировать — она не опровергает принцип независимости: эти объединения можно воспринимать как одно целое. И, в конце-то концов! — такова принятая нами модель нервных систем, и она, как и любая другая, имеет право на некоторое несоответствие, лишь бы была полезной и плодотворной.
Замыкает цепочку от рецептора до мышцы синапс коры головного мозга — самое узкое место канала, выполняющее функцию регулируемого дросселя. Самой главной его особенностью является то, что он может изменять свою проводимость — проходное сечение — в широких пределах. Такое свойство присуще исключительно только синапсам коры головного мозга; другие синапсы, соединяющие нейроны и мотонейроны в цепочки, не способны изменять свою проводимость, и по этой причине им лучше бы дать иное название. Свою проводимость синапс изменяет в процессе обучения; обученные синапсы определяют те законы, которые увязывают однозначно ситуации с поведением объекта в них. Таким образом, саморегулируемый синапс является главным элементом нервных систем животных: он представляет собой как раз то «конструктивное» отличие, с помощью которого Природа превратила системы безусловных рефлексов в системы условных, в системы навыков. Без преувеличения можно сказать, что появление синапса коры головного мозга, точнее — его регулируемых свойств, породило весь животный мир; такое изобретение Природы стоит очень и очень дорого.
Итак,
конкретный синапс встроен в канал управления, соединяющий конкретный рецептор
с конкретной мышцей; каждый рецепторный нейрон, таким образом, имеет
синаптическую связь со всеми мотонейронами
и, наоборот, каждый мотонейрон имеет такую же связь со всеми рецепторными
нейронами. В результате структура коры приобретает матричный вид со строками и
столбцами; условимся нейроны представлять как строки, а мотонейроны — как столбцы матрицы, на пересечении тех и
других — соединяющие их синапсы.
Математическая модель работы нервных
систем. Рассмотрим систему с полным набором рецепторов, общее количество
которых пусть будет m,
и с одним мотонейроном. Такая система может управлять только одним
исполнительным органом; для любого другого исполнительного органа потребуется
точно такая же система, и она может быть объединена с первой, не мешая ей, в
соответствии о принципом независимости. Обозначим проводимость i-го
синапса, соответствующего i-му рецептору, через ![]() . Принимая течение управляющей жидкости в синапсе ламинарным,
можем получить выражение для определения потока, проходящего через синапс как
произведение возбуждения рецептора
. Принимая течение управляющей жидкости в синапсе ламинарным,
можем получить выражение для определения потока, проходящего через синапс как
произведение возбуждения рецептора ![]() на проводимость
соответствующего синапса
на проводимость
соответствующего синапса ![]() :
:
![]() .
(1.1)
.
(1.1)
Этот поток можно считать рецепторной долей общего сигнала управления; полная величина этого сигнала определится как
![]() . (1.2)
. (1.2)
Выражения (1.1) и (1.2)
отражают работу системы навыков и условных рефлексов животных (нервной
системы) или, что одно и тоже, обучаемой системы управления. На первый взгляд
указанные выражения относят эти системы управления в разряд простейших линейных,
в то время как реальные зависимости сигналов управления какого-то бы ни было
объекта от внешних параметров, как мы знаем, всегда
нелинейны; но это — только на первый
взгляд. Проблема линейности-нелинейности будет подробно рассмотрена ниже, а
здесь отметим лишь, что системы навыков и условных рефлексов работают не с
внешними параметрами, а с внутренними образами очувствления; очувствление
объекта может быть дискретизировано (разбито) на
столь малые величины, то есть количество рецепторов можно выбрать таким
большим, что в пределе система управления превратится в непрерывную, и
рассуждения о линейности-нелинейности потеряют смысл. Выражение (1.1) принято
линейным только из соображений простоты, а в самом деле оно может быть каким
угодно нелинейным при условии сохранения прямой зависимости рецепторной доли ![]() от возбуждения
от возбуждения ![]() и проводимости
и проводимости ![]() ; и, наконец, обучаемые
системы управления не являются логическими со строгим математическим
обеспечением, и выдают сигналы управления, можно сказать, всегда неточными, но в пределах допустимых отклонений.
; и, наконец, обучаемые
системы управления не являются логическими со строгим математическим
обеспечением, и выдают сигналы управления, можно сказать, всегда неточными, но в пределах допустимых отклонений.
Выше мы
упоминали, что электрический биоток является всего лишь сопутствующим явлением и используется только при воздействии на
мышцы, однако электрический потенциал действия возникает не в конце
мотонейрона, а в самом начале нейронов. Почему так? Можно предположить, что
электрический потенциал действия способствует поддержанию волны давления;
иначе она в вязкой жидкости очень скоро могла бы погаснуть. Как только на
переднем фронте волны вскроются поры,
через них под воздействием электрического притяжения устремляются внутрь ионы
натрия; они-то и подпитывают волну. На гребне волны и на обратном фронте ионы
натрия выдавливаются обратно.
И ещё один
вопрос: почему могут бездействовать мышцы животного в состоянии покоя? Можно
сослаться, конечно, на пониженный тонус организма, но это — не главное. Нужно учесть ещё то, что
процессу напряжения мышцы всегда сопутствует процесс расслабления: положительно
заряженные порции управляющей жидкости, поступающие на мышцы, непрерывно
нейтрализуются питательной средой. В гидравлической схеме это равносильно
утечкам. Сокращение мышц может возникнуть только в том случае, если поток
управляющей жидкости превысит нейтрализацию. Таков в общих чертах механизм
нервных систем животных и систем, реализующих навыки и условные рефлексы
человека; таков же механизм и обучаемых систем управления, хотя некоторые
особенности они имеют.
Электрическая обучаемая система управления. Выше говорилось, что по виду энергии обучаемые системы могут быть гидравлическими, пневматическими, оптическими и, конечно, электрическими. Последнюю систему, как наиболее характерную, рассмотрим более подробно. На рис.1.8. представлена простейшая аналоговая схема с выходом на один исполнительный орган; при необходимости число выходов может быть увеличено, и при этом структура системы сохранится прежней.
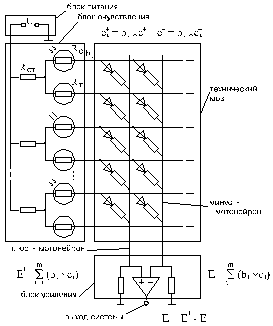
Рис.1.8. Принципиальная схема электрической аналоговой обучаемой обучаемой системы управления на один исполнительный орган
Основным
отличием технической обучаемой системы от нервной системы животных является
то, что она должна выдавать на выходе двуполярный сигнал управления, то есть и
плюс, и минус, потому что, как правило, все двигатели имеют и прямое, и реверсивное
(обратное) движения; в то время как мышцы могут только тянуть и не способны
толкать, и поэтому для них достаточно иметь сигнал управления только одного
знака. Для реализации двухсторонних движений исполнительных органов и получения
двуполярного сигнала управления в систему введены по два столбца матрицы мозга
на каждый отдельный исполнительный орган: один столбец формирует положительный
сигнал управления, а другой — отрицательный; результирующий сигнал определяется
сравнением (вычитанием отрицательного сигнала из положительного).
Структура
системы (рис. 1.8) представлена в блочном
виде; штриховыми линиями выделены блоки: питания, очувствления, усиления, — и
блок, представляющий технический мозг. Питание осуществляется от источника
постоянного тока неограниченной мощности (
U — напряжение питания): таким образом, рецепторы технической
системы, в отличие от рецепторов
животных, не иссякают и не нуждаются в подзарядке, то есть во сне. Сами
рецепторы представляют собой резисторы; рецепторы спарены: на один рецептор
прямого действия, возбуждающийся при
внешнем воздействии на него, приходится один рецептор обратного действия,
электрическое напряжение на выходе которого растет при снижении внешнего
воздействия на первый.
Поясним это на
примере. Допустим, фоторецептором прямого действия является фоторезистор,
сопротивление которого в зависимости от освещённости изменяется в пределах ![]() =
20...2000 кОм. Сопротивления двух других резисторов рецепторной пары
таковы: спаренного резистора
=
20...2000 кОм. Сопротивления двух других резисторов рецепторной пары
таковы: спаренного резистора ![]() = 200 кОм и стабилизирующего резистора, стоящего на входе
пары,
= 200 кОм и стабилизирующего резистора, стоящего на входе
пары, ![]() = 2 000 кОм. Тогда при яркой освещённости:
= 2 000 кОм. Тогда при яркой освещённости: ![]() = 20 кОм;
= 20 кОм; ![]() =10
=10![]() , где
, где ![]() — ток, проходящий
через фоторезистор, а
— ток, проходящий
через фоторезистор, а ![]() — ток, проходящий
через спаренный резистор. При средней освещённости:
— ток, проходящий
через спаренный резистор. При средней освещённости: ![]() = 200 кОм;
= 200 кОм; ![]() =
=![]() ; ив темноте:
; ив темноте: ![]() = 2000 кОм;
= 2000 кОм; ![]() =0,1
=0,1![]() . Таким образом, в каждой отдельной паре
рецепторов при сильном внешнем воздействии больше возбуждён первый их них, и
поэтому его можно считать рецептором прямого действия, а при слабом — второй, и он является рецептором обратного
действия.
. Таким образом, в каждой отдельной паре
рецепторов при сильном внешнем воздействии больше возбуждён первый их них, и
поэтому его можно считать рецептором прямого действия, а при слабом — второй, и он является рецептором обратного
действия.
В качестве технических органов чувств, составляющих блок очувствления могут быть фоторезисторы, реагирующие на освещенность, терморезиоторы, реагирующие на теплоту, силарезиоторы, например из токопроводящей резины, реагирующие на усилие нажатия, звукорезисторы, реагирующие на силу звука в узком диапазоне частот и другие. Рецепторы очувствления образуют рецепторные поля, например сетчатку глаза или чувствительную техническую кожу; количество рецепторов в рецепторном поле не ограничено; расположение рецепторов в рецепторном поле — произвольное.
Технический мозг представляет собой матрицу, имеющую строчные и столбцовые шины, на пересечении которых установлены аналоговые резисторные элементы — синапсы. К строчным шинам подсоединены выходы рецепторов; столбцовые шины (мотонейроны) спарены: одну из них назовем плюс-столбцом (плюс-мотонейроном), а вторую — минус-столбцом (минус-мотонейроном). Аналоговый резисторный элемент мозга — синапс — может представлять собой подстроечный или иной регулируемый вручную резистор. Также в качестве аналоговых резисторных элементов могут использоваться такие регулируемые резисторы, изменение сопротивления которых изменяется автоматически под воздействием общих команд, подаваемых обучателем, и в зависимости от возбуждения соответствующих рецепторов. Такими резисторами могут быть, в частности, аморфные полупроводники, например из халькогенидного отекла. Для того, чтобы исключить появление в матрице технического мозга блуждающих токов, в каждую цепь с аналоговым резисторным элементом встроен диод, пропускавший ток только в одном направлении. Необходимость в диодах отпадает при условии, что соотношение сопротивлений рецепторов очувствления, аналоговых резисторных элементов мозга и прочих резисторов цепи обеспечивает только одно заданное направление тока. Необходимость в них отпадает также и в том случае, если используется ток переменного направления.
Блок усиления включает нагрузочные резисторы и усилители; нагрузочные резисторы соединяют столбцовые шины мозга с источником питания; усилители совмещают функции сумматоров-вычитателей и усилителей сигналов управления. Каждая пара столбцов подсоединена к своему усилителю: плюс-столбец подсоединён к неинверсному (+) входу, а минус-столбец — к инверсному (-). Выход усилителя является выходом всей системы на отдельный двигатель, а степень усиления усилителя подбирается такой, чтобы сигнал управления на выходе имел стандартный диапазон.
Пользуясь
случаем отметим, что в двуполярной
обучаемой системе управления, у которой сигнал на выходе может менять свой
знак, наблюдается полная 'симметрия: плюс- и минус-столбцы матрицы мозга
абсолютно равноправны и равноценны, и обозначение их плюсом и минусом условно;
поэтому было бы совершенно несправедливо по отношению к сигналам управления (и,
в конце концов, к исполнительным органам) считать, что, например, сигнал ![]() = +5 мВ больше сигнала
= +5 мВ больше сигнала ![]() = -5 мВ: в данном случае они равны, но
противостоят друг другу. Приходится сожалеть, что нет такой символики, кроме (+)
и (-), которая обозначала бы
противостоящие величины.
= -5 мВ: в данном случае они равны, но
противостоят друг другу. Приходится сожалеть, что нет такой символики, кроме (+)
и (-), которая обозначала бы
противостоящие величины.
Работа обучаемой системы управления. В
исходном состоянии проводимости всех синапсов мозга равны нулю или, точнее сказать,
наименьшие возможные, так что, если и возникает на спаренных столбцовых шинах электрическое напряжение, то оно —
одинаковое и на плюс-, и на минус-столбцах, а на выходе усилителя — ноль. В процессе
обучения с многократными повторами проводимости синапсов будут изменяться так
долго, пока не завершится обучение, и в результате они установятся вполне определёнными
и у каждого синапса — своя: ![]() — проводимость синапса, связанного с i-ым рецептором и с
плюс-столбцом матрицы мозга,
— проводимость синапса, связанного с i-ым рецептором и с
плюс-столбцом матрицы мозга, ![]() — то же — с минус-столбцом.
— то же — с минус-столбцом.
В обученном
рабочем состоянии обучаемая система выдает на выходе такой сигнал управления,
который соответствует текущей ситуации. Как это происходит? Любая внешняя j-ая ситуация порождает
определенный образ внутреннего очувствления системы; это означает, что в этой
ситуации каждый i-ый рецептор будет
иметь свое конкретное возбуждение ![]() и с учетом соответствующих
ему проводимостей
и с учетом соответствующих
ему проводимостей ![]() и
и ![]() выдаст в плюс-столбец
матрицы мозга такую свою рецепторную долю сигнала управления:
выдаст в плюс-столбец
матрицы мозга такую свою рецепторную долю сигнала управления:
![]() ,
(1.3)
,
(1.3)
а в минус-столбец - такую:
![]() ,
(1.4)
,
(1.4)
В результате суммирования долей (1.3) всех рецепторов на плюс-столбце матрицы определится общий положительный сигнал управления, известный в нейрофизиологии как возбуждение:
![]() , (1.5)
, (1.5)
а суммирование долей (1.4) на минус-столбце даст общий отрицательный сигнал управления ( торможение):
![]() , (1.6)
, (1.6)
и результирующий сигнал управления системы, вычисляемый сумматором-вычитателем (усилителем, выполняющим функции элемента сравнения) , определится как
![]() . (1.7)
. (1.7)
Выражения (1.1)...(1.7) отражают работу обучаемых систем управления, а точнее — только одного ее исполнительного органа; однако, согласно принципа независимости, эти выражения в равной степени относятся и к другим органам,
Для того,
чтобы глубже понять обучаемые системы управления, воспользуемся образным
представлением и изобразим действия, связанные с этими системами в виде
|
|
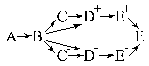
(1.8)
где А — внешняя ситуация; В —
образ внутреннего очувствления, то есть образ возбуждений рецепторов; ![]() — образ проводимостей синапсов
плюс-мотонейрона;
— образ проводимостей синапсов
плюс-мотонейрона; ![]() — то же самое
минус-мотонейрона ;
— то же самое
минус-мотонейрона ; ![]() — образ
рецепторных долей плюс-мотонейрона;
— образ
рецепторных долей плюс-мотонейрона; ![]() — образ рецепторных
долей минус-мотонейрона;
— образ рецепторных
долей минус-мотонейрона; ![]() ,
, ![]() ;
; ![]() — скалярные величины сигналов управления,
формируемые соответственно на плюс-мотонейроне, на минус-мотонейроне и на
выходе системы. В качестве примечания отметим, что усиление сигнала управления
на выходе преследует цель лишь изменить масштаб сигнала и никак не меняет его зависимость от ситуаций.
— скалярные величины сигналов управления,
формируемые соответственно на плюс-мотонейроне, на минус-мотонейроне и на
выходе системы. В качестве примечания отметим, что усиление сигнала управления
на выходе преследует цель лишь изменить масштаб сигнала и никак не меняет его зависимость от ситуаций.
Дадим некоторые пояснения к выражению (1.8). Система очувствления реализует следующую зависимость
![]() ,
,
и это можно истолковать как преобразование внешней ситуации во внутренний образ; в этом преобразовании рецепторное поле выступает в качестве преобразующей среды, проходя через которую параллельным потоком, внешняя информация превращается в параллельные потоки внутренней.
В результате обучения под воздействием внутреннего образа В формируются образы проводимостей синапсов мозга:
при поощрении:
![]() ;
;
при наказании:
![]() .
.
Образы ![]() и
и ![]() возникают уже как
наложение образа В на образы
возникают уже как
наложение образа В на образы ![]() и
и ![]() :
:
![]()
Если
представить образ В в виде
зрительного изображения, собранного оптикой, а образы ![]() и
и ![]() в виде некоторых
изображений на диапозитивах, то образы
в виде некоторых
изображений на диапозитивах, то образы ![]() и
и ![]() возникнут при
пропускании изображений В через
указанные диапозитивы.
возникнут при
пропускании изображений В через
указанные диапозитивы.
![]()
то есть в примере с пропусканием зрительного изображения через диапозитивы операции суммирования представляют собой определение суммарных световых потоков на выходах из диапозитивов.
И, наконец, выполняется простая операция вычитания:
![]() .
.
Таков в общих
чертах механизм обучаемых систем управления: ее устройство и ее работа.
Осталось только пояснить, что такое — возбуждающие и тормозящие рецепторы.
Разделение рецепторов на возбуждающие и тормозящие — не врождённое, а происходит в результате обучения. Если
окажется после завершения обучения, что ![]() , то соответствующий рецептор можно считать возбуждающим,
так как его доля в общем сигнале управления окажется положительной:
, то соответствующий рецептор можно считать возбуждающим,
так как его доля в общем сигнале управления окажется положительной:
![]() ,
,
и, наоборот, если окажется, что ![]() , то соответствующий рецептор можно считать тормозящим, так
как его доля окажется отрицательной:
, то соответствующий рецептор можно считать тормозящим, так
как его доля окажется отрицательной:
![]()